

Signal peptides are short amino acid sequences with an outsized effect on where proteins go and what they become. A newly made polypeptide may be destined for secretion, insertion into a membrane, or residence within the secretory system, and a signal peptide is often the first instruction that sets that path.
The term can sound simple at first glance. In practice, it covers several related sequence classes, multiple trafficking routes, distinct cleavage outcomes, and even a separate cosmetic meaning that has little to do with intracellular protein targeting. Keeping those uses apart makes the biology much clearer.
What signal peptides are in protein targeting
In cell biology, a signal peptide is usually a short targeting sequence near the amino terminus of a protein. Its job is not to supply catalytic activity or structural stability. Its job is to tell the cell, early, where the nascent protein should go.
Many classical signal peptides are about 15 to 30 amino acids long and contain a prominent hydrophobic stretch near the N terminus. Even so, there is no single universal consensus sequence. Signal peptides are recognized more by their overall physicochemical pattern than by one exact motif. That is why cells can route a wide range of proteins through the same general machinery.
A useful way to think about signal peptides is as modular sequence elements with a recurring three-part architecture:
- N-region: short segment near the amino terminus, often enriched in positively charged residues
- H-region: hydrophobic core that interacts with recognition factors and membrane translocation machinery
- C-region: segment near the cleavage site, often containing small neutral residues favored by signal peptidase
That architecture is common, not mandatory. Biological systems tolerate variation, and pathway context matters.
Eukaryotic ER targeting pathways for signal peptides
In eukaryotic cells, the best-known destination for a classical signal peptide is the rough endoplasmic reticulum. Secreted proteins, many lysosomal proteins, and many membrane proteins enter the ER early in synthesis and then continue through the secretory pathway.
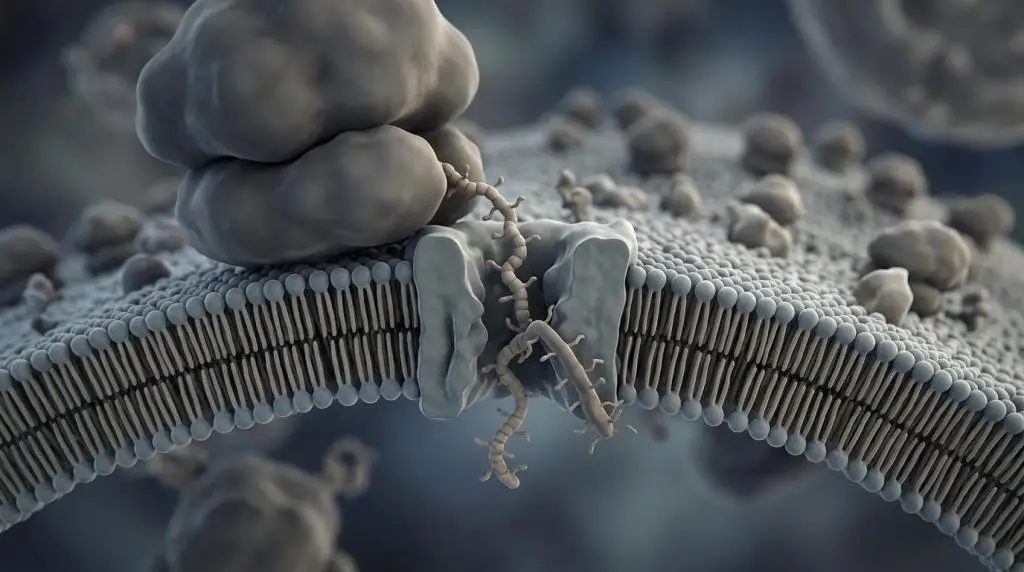
The standard route is co-translational translocation. As the signal peptide emerges from the ribosome, it is recognized by the signal recognition particle, or SRP. SRP temporarily slows translation, guides the ribosome nascent-chain complex to the ER membrane, and transfers it to the SRP receptor. The ribosome then engages the Sec61 translocon, a protein-conducting channel in the ER membrane.
From there, several outcomes are possible. A soluble secretory protein may pass through the channel into the ER lumen, where the signal peptide is commonly cleaved by signal peptidase. A membrane protein may instead use the same early targeting logic but become embedded in the membrane through signal-anchor or stop-transfer segments that define topology.
The core steps are often summarized this way:
- Signal peptide emerges from the ribosome.
- SRP binds the signal peptide and ribosome.
- The complex docks at the rough ER through the SRP receptor and Sec61.
- Translation resumes, driving translocation into or across the membrane.
Not every ER-directed protein depends on SRP in exactly the same way. Eukaryotic cells also support post-translational import into the ER, especially for some smaller or less hydrophobic substrates. In that route, Sec61 still serves as the translocation channel, but accessory factors including the Sec62/63 complex and the luminal chaperone BiP help pull an unfolded chain through the membrane. That detail matters because it shows that a “signal peptide” is not tied to one single delivery model.
Bacterial secretion pathways and bacterial signal peptides
Bacteria use related logic with different hardware. Proteins that must cross or insert into the plasma membrane often begin with an N-terminal signal peptide that is read by secretion or membrane-targeting systems. The best known is the Sec pathway, which translocates proteins in an unfolded state.
Bacterial signal peptides also tend to show an N-region, a hydrophobic core, and a cleavage region. In many cases, the N-region carries positive charge, the hydrophobic region drives membrane engagement, and the C-region positions the peptide for cleavage by signal peptidase after export begins or completes. The overall principle is familiar even though the machinery is not identical to the eukaryotic ER system.
Bacteria also use SRP. In many species, the SRP protein Ffh recognizes hydrophobic signal sequences as they emerge from the ribosome and helps target nascent membrane proteins to the membrane. That means SRP is not exclusively a eukaryotic story. It is a conserved solution to the problem of routing hydrophobic proteins before they aggregate in the cytosol.
A second major bacterial route is the twin-arginine translocation, or Tat, pathway. Tat signal peptides carry a hallmark twin-arginine motif and direct proteins across the membrane in a folded state. This is a major contrast with the Sec pathway, which usually handles unfolded substrates.
| Pathway | Typical location | Signal sequence feature | Timing of targeting | Common outcome |
|---|---|---|---|---|
| SRP to Sec61 | Eukaryotic rough ER | Hydrophobic N-terminal signal peptide or signal-anchor | Usually co-translational | Secretion into ER lumen or membrane insertion |
| Sec62/63 to Sec61 | Eukaryotic rough ER | ER-targeting signal peptide | Post-translational | Import of selected unfolded proteins into ER |
| Sec pathway | Bacterial plasma membrane | Classical bacterial signal peptide | Co-translational or post-translational, depending on substrate | Export or membrane insertion of unfolded proteins |
| Tat pathway | Bacterial plasma membrane | Signal peptide with twin-arginine motif | Post-translational | Export of folded proteins |
Types of signal peptides and membrane-targeting sequences
Signal peptides are not one uniform class. A better classification starts with three questions: where the sequence is located, whether it is cleaved, and which pathway recognizes it.
The cleavable N-terminal signal peptide is the textbook example. It initiates targeting to a translocation system and is then removed by signal peptidase. Many secreted proteins use this design. Once cleavage occurs, the mature protein no longer carries the original targeting tag.
Noncleavable signal sequences are different. A signal-anchor sequence can both target the ribosome nascent-chain complex to the membrane and remain in the final protein as a transmembrane helix. Internal signal sequences can do the same job, especially in membrane proteins that do not begin with a classical cleavable signal peptide.
This is where membrane protein biogenesis becomes more interesting.
A single hydrophobic segment can act first as a targeting cue and later as part of the mature protein’s topology. Additional hydrophobic segments may function as start-transfer or stop-transfer sequences, setting the orientation of loops and domains on either side of the membrane. Multipass membrane proteins rely on this repeated logic to build a stable arrangement across the bilayer.
That is why “signal peptide” is often used loosely in casual discussion, while more precise writing separates cleavable signal peptides from signal anchors, internal signal sequences, and other topogenic elements.
What controls signal peptide cleavage and membrane topology
Cleavage is not automatic. Signal peptidase recognizes a sequence context near the C-region of a cleavable signal peptide, and small residues close to the cleavage site are often favored. If the sequence is highly hydrophobic in the wrong way, placed differently within the chain, or constrained by neighboring structure, cleavage may not occur.
Orientation also depends on more than one feature. The distribution of positive charges around a transmembrane segment, often described by the positive-inside rule, can bias which side of the membrane remains cytosolic. Hydrophobic segment length, the timing of emergence from the ribosome, chaperone contacts, and neighboring transmembrane helices all shape the final topology.
This has direct experimental value. A point mutation that weakens hydrophobicity or alters residues near the cleavage site can redirect trafficking, reduce secretion efficiency, or invert membrane orientation. In protein engineering, signal peptide choice is often one of the first variables adjusted when a construct expresses poorly or mislocalizes.
Signal peptides in skincare use a different definition
Outside cell biology, the phrase signal peptide also appears in skincare. Here, the meaning shifts. Cosmetic “signal peptides” are short bioactive peptides marketed for their ability to stimulate skin cells to produce matrix components or respond in a repair-associated way. They are not the same thing as the N-terminal targeting sequences that send proteins to the ER, bacterial membrane, or secretion pathways.
That distinction matters because the two uses share a name but not a mechanism. A cellular signal peptide is part of a larger protein and functions as an intracellular address label during biosynthesis. A cosmetic signal peptide is a standalone active ingredient category used in topical formulations.
Topical peptide products are often grouped into a few broad categories:
- signal peptides
- Carrier peptides: peptides intended to deliver or bind ions, often metals, in skin-focused formulations
- Neurotransmitter-inhibiting peptides: peptides marketed to influence signaling linked to expression lines
- Enzyme-inhibiting peptides: peptides positioned as modulators of enzymatic processes tied to skin appearance
When a paper, product page, or label uses the term, context is everything. If the discussion mentions SRP, Sec61, rough ER, secretion, translocation, or cleavage by signal peptidase, it is using the cell biology meaning. If it mentions fibroblasts, collagen support, topical application, or anti-aging categories, it is using the cosmetic one.
How to read signal peptide annotations in papers and databases
In research articles and sequence databases, “signal peptide” usually signals a prediction about protein trafficking. Annotation tools look for the characteristic N-terminal pattern, hydrophobicity profile, and probable cleavage site. That annotation often implies that the mature protein enters the secretory pathway, though it does not by itself prove the final subcellular location.
A good reading habit is to pair the annotation with other clues. Is there a transmembrane helix after the signal peptide? Is the protein secreted or membrane-bound in experimental data? Does the sequence contain a twin-arginine motif, suggesting Tat export in bacteria? Does the paper describe SRP dependence or Sec62/63 involvement? Those details turn a generic label into a specific trafficking model.
For researchers and advanced readers, precision here pays off quickly. It sharpens construct design, improves interpretation of localization data, and prevents confusion when the same term appears in molecular cell biology and skincare marketing. Signal peptides may be short, but they sit at the center of one of the cell’s most elegant sorting systems.